Fajképződés (speciáció): meghatározás, mechanizmusok és példák
Fajképződés (speciáció): áttekintés — meghatározás, izolációs és hibridizációs mechanizmusok, folyamatok és szemléletes példák evolúcióbiológiai kontextusban.
A fajképződés arról szól, hogyan alakulnak ki a fajok. Ez az evolúcióbiológia egyik legfontosabb része.
Darwin úgy gondolta, hogy a legtöbb faj közvetlenül a már létező fajokból alakult ki. Ezt nevezik anagenezisnek: a fajok változás útján történő kialakulása, vagy más néven "phyletikus evolúció". A 20. század nagy részében úgy gondoltuk, hogy a legtöbb faj korábbi fajok szétválásával jött létre: kladogenezis. Az általános nézet az volt, hogy a legtöbb fajfelosztást izoláló mechanizmusok okozzák vagy segítik útjukat.
Kétségtelen, hogy az egykor együtt élő fajok fizikai szétválása az egyik fő tényező. Ezt nagyon sok példa szemlélteti, amelyek közül néhányat az alábbiakban tárgyalunk.
Az elmúlt 20 évben végzett munka azonban más okokat is kimutatott. Az élőlények DNS-szekvenciájának elemzése kimutatta, hogy a rokon fajok között gyakran van némi hibridizáció. Ez azt jelenti, hogy ezek a kereszteződések géneket adtak át. Ez viszont azt jelenti, hogy nem a reproduktív izoláció a faj egyetlen definíciója, és a fajképződéshez nem mindig van szükség allopatriára (a fajok szaporodási szempontból elkülönülnek egymástól). Az alábbi szakaszok azt az elképzelést illusztrálják, hogy a fizikai elkülönülés elsődleges fontosságú volt az új fajok kialakulásában.
Képgaléria
10 Képek









Mi az a fajképződés?
A fajképződés (speciáció) olyan folyamat, amely során egy vagy több új, egymástól reproduktívan elkülönült faj jön létre egy vagy több elődpopulációból. A folyamat lehet lassú, fokozatos változás eredménye, de történhet viszonylag rövid idő alatt is, ha például nagy genetikai változások (pl. poliploidizáció) következnek be.
Fő mechanizmusok
A fajképződés lehetséges útjai közül a legfontosabbak:
- Allopatrikus speciáció: a populációk földrajzi elkülönülése (pl. hegység, folyó kialakulása) megakadályozza a génáramlást. Idővel különböző mutációk és szelekciós nyomások eltérő irányba viszik a genetikát, ami reproduktív izolációhoz vezet.
- Peripatrikus speciáció: egy kis, elszigetelt populáció alapítói effektus és erős genetikai sodródás hatására gyorsabban diverzifikálódhat az anyaállománytól.
- Parapatrikus speciáció: szomszédos populációk egymáshoz viszonyított részleges izolációja esetén, ahol a környezeti gradiens mentén erős lokális szelekció és csökkent génáramlás egyaránt szerepet játszik.
- Szimpátrikus speciáció: ugyanazon földrajzi területen belül alakul ki az új faj, például ha különböző forrástípusokra történő táplálkozási specializáció vagy szexuális szelekció osztja fel a populációt. A növényeknél tipikus példa a poliploidizáció, amikor duplikálódik a kromoszómaszám, és az így keletkező példány már rögtön reproduktívan elkülönülhet az anyaformától.
Elkülönülést biztosító mechanizmusok
Az izoláció lehet prezygotikus (megakadályozza a párosodást vagy a megtermékenyítést) és postzygotikus (a hibrid utódok csökkent életképessége vagy terméketlensége). Példák:
- Időbeli izoláció (eltérő szaporodási időszakok)
- Viselkedési izoláció (eltérő párválasztási jelzések vagy szokások)
- Mechanikai izoláció (társuló morfológiai eltérések)
- Gamétikus izoláció (ivarsejtek között nem jön létre sikeres megtermékenyítés)
- Hibrid csökkent életképessége vagy terméketlensége (postzygotikus)
Genetikai alapok és fontos fogalmak
A fajképződés genetikai hátterében gyakran Dobzhansky–Muller típusú inkompatibilitások állnak: két populáció különböző mutációkat halmoz fel, amelyek együtt rosszul működnek, amikor újra találkoznak. Emellett a lokalizált adaptáció, a szelekció, a genetikai sodródás és a génáramlás (gene flow) együttesen határozzák meg, hogy izoláció kialakul-e.
Reinforcement akkor történhet, amikor a hibridizáció költséges (hibrid utódok kevésbé életképesek), és a természetes kiválasztódás fokozza a prezygotikus izolációt, hogy elkerülje a nem sikeres kereszteződéseket.
Példák a természetből
- Darwin-pintyek: a Galápagos-szigeteken élő pintyek példája jól mutatja az allopatrikus és adaptív radiáció szerepét, ahol különböző szigetek és táplálkozási források különböző fajok kialakulásához vezettek.
- Rhagoletis alma-legyek: az alma és vadalma (hawthorn) táplálkozási specializációja klasszikus példája a szimpátrikus speciációnak, ahol táplálékpreferencia és időbeli reprodukciós különbségek hoztak létre reproduktív elkülönülést.
- Halak, például afrikai tavi cichlidek: gyors adaptív radiáció és szexuális szelekció miatt rendkívül sok faj alakult ki rövid földtörténeti idő alatt, gyakran kis geológiai területeken belül.
- Növények poliploidizációja: sok kultúrfaj (pl. búza egyes variánsai) és vadnövény esetében a kromoszómaszám duplikációja azonnali reproduktív izolációt eredményezhet.
Hibridizáció és genomika szerepe
Az utóbbi évtizedekben a genomikai vizsgálatok kimutatták, hogy a hibridizáció és a introgresszió (idegen gének beépülése) gyakori, és sokszor hozzájárul a helyi adaptációhoz. Ez azt jelenti, hogy a fajhatárok nem mindig élesek: bizonyos gének átjárhatnak fajok között, miközben más genomi régiók erősen elkülönülnek.
Mennyi idő alatt alakul ki egy faj?
A válasz nagyon változó: egyes esetekben néhány generáció (poliploid növények) is elég, máskor több százezer vagy millió év a tipikus időskála. A sebességet befolyásolja a populációméret, a szelekció ereje, a környezeti változások és a génáramlás mértéke.
Összefoglalás
A fajképződés sokrétű, többféle mechanizmus együttműködéséből kialakuló folyamat. Bár a földrajzi elkülönülés (allopatria) gyakran fontos, nem ez az egyetlen út: szimpátrikus, parapatrikus és peripatrikus mechanizmusok, poliploidizáció, hibridizáció és lokális adaptáció is létrehozhatnak új fajokat. A modern genomikai eszközökkel egyre részletesebben érthetjük meg, hogyan formálódnak a fajok és milyen szerepet játszanak a különböző genetikai és ökológiai tényezők a speciáció folyamatában.
Elszigetelő mechanizmusok
Az elkülönítő mechanizmusok olyan dolgok, amelyek megakadályozzák a sikeres szaporodást egy faj csoportjai között. A populációk szaporodási izolációja létrejön. Ez különösen fontos a biológiai fajfogalom szempontjából, mivel a fajokat a szaporodási izoláció határozza meg.
Az izoláló mechanizmusok két csoportra oszthatók, a megtermékenyítés előtti és utáni csoportra.
Megtermékenyítés előtt
Olyan tényezők, amelyek megakadályozzák az egyedek párosodását.
- Földrajzi elszigeteltség: A fajok különböző területeken fordulnak elő, és gyakran akadályok választják el őket egymástól.
- Időbeli elszigeteltség: Az egyedek nem párosodnak, mert különböző időpontokban aktívak. Ez lehet különböző napszakok vagy különböző évszakok. Előfordulhat, hogy a fajok párzási időszakai nem egyeznek meg. Az egyedek nem találkoznak egymással sem a párzási időszakuk alatt, sem egyáltalán.
- Ökológiai elszigeteltség: Az egyedek csak az általuk preferált élőhelyen párosodnak. Nem találkoznak más fajok más ökológiai preferenciájú egyedeivel.
- Viselkedésbeli elszigeteltség: A különböző fajok egyedei találkozhatnak, de az egyik nem ismeri fel az esetlegesen adott szexuális jeleket. Az egyed a legtöbb esetben a saját fajának egy tagját választja.
- Mechanikai szigetelés: Megkísérelhető a kopuláció, de a spermiumok átvitelére nem kerül sor. Az egyedek mérete vagy morfológiája miatt összeférhetetlenek lehetnek.
- Gametikus inkompatibilitás: A spermaátvitel megtörténik, de a petesejt nem termékenyül meg.
Megtermékenyítés után
Olyan tényezők, amelyek megakadályozzák a párosodás sikerét, például genetikai inkompatibilitás, hibrid életképtelenség vagy sterilitás.
- Zigóta mortalitás: A petesejt megtermékenyül, de a zigóta nem fejlődik ki.
- Hibrid életképtelenség: Hibrid embrió alakul ki, de nem életképes.
- Hibrid sterilitás: A hibrid életképes, de az ebből származó felnőtt egyed steril.
- Hibrid bontás: Az első generációs (F1) hibridek életképesek és termékenyek, de a további hibridgenerációk (F2 és visszakeresztezések) életképtelenek vagy sterilek.

Földrajzi elszigeteltség
Ezt tartják a fajképződés leggyakoribb okának. Elsőként Moritz Wagner német felfedező és természettudós jutott erre az ötletre.
Wagner pályája elején geográfusként dolgozott, és számos földrajzi könyvet publikált Észak-Afrikáról, a Közel-Keletről és a trópusi Amerikáról. Emellett lelkes természettudós és gyűjtő is volt, és a biológusok körében leginkább erről a munkájáról ismert. Ernst Mayr, az evolucionista és biológiatörténész számolt be Wagner jelentőségéről. p562-565
Algériában töltött három éve alatt Wagner (egyéb tevékenységei mellett) a Pimelia és Melasoma nevű röpképtelen bogarakat tanulmányozta. Mindkét nemzetség több fajra oszlik, amelyek mindegyike az Atlasz-hegységből a Földközi-tengerbe ereszkedő folyók közötti északi partvidék egy-egy szakaszára korlátozódik. Amint az egyik átkel egy folyón, egy másik, de közeli rokon faj jelenik meg.
"... egy kezdődő faj csak akkor [keletkezik], ha néhány egyed átlépi elterjedési területének határait... egy új faj kialakulása soha nem fog sikerülni... anélkül, hogy a telepesek hosszú időn át ne különülnének el fajuk többi tagjától".
Ez egy korai leírása volt egyfajta földrajzi fajképződési folyamatnak. Mayr 1942-ben újra bevezette, és a földrajzi speciáció fontossága az evolúciós szintézis egyik központi gondolatává vált.
A földrajzi fajképződés másik kifejezése az allopatrikus fajképződés. Az allopatria azt jelenti, hogy "különböző föld".
Üres szigetek
A vulkanikus szigetek élet nélkül keletkeznek, és minden életnek szél vagy víz által kell érkeznie. A Hawaii-szigetekről és a Galapagos-szigetekről tudjuk, hogy minden életforma megváltozik, amikor a szárazföldről a szigetekre érkezik.
A Hawaii-szigeteken, mintegy 6 500 négyzetkilométeren (17 000 km2) található a világ legváltozatosabb drosophilid légygyűjteménye, amely az esőerdőtől a hegyi rétekig él. Mintegy 800 hawaii drosophilid faj ismert.
A tanulmányok a fajok egyértelmű "áramlását" mutatják a régebbi szigetekről az újabbak felé. Vannak esetek a régebbi szigetekre való visszatelepülésre és a szigetek kihagyására is, de ezek sokkal ritkábbak.
A kálium/argon radioaktív kormeghatározás szerint a jelenlegi szigetek 0,4 millió évvel ezelőtt (mya) (Mauna Kea) és 10mya (Necker) közöttiek. A hawaii szigetvilág legrégebbi, még a tenger felett lévő tagja a Kure-atoll, amely 30 mya-re datálható.
Maga a szigetcsoport, amelyet a Csendes-óceáni lemez egy forró ponton való mozgása hozott létre, sokkal régebb óta létezik, legalábbis a kréta időszakig. A Hawaii-szigetek és a korábbi szigetek, amelyek most a tenger alatt vannak, alkotják a Hawaii-császár-tengeri hegyláncot; és a víz alatti hegyek közül sokan srácok.
Úgy tűnik, hogy a Hawaiiban honos összes drosophilid faj egyetlen ősi fajból származik, amely körülbelül 20 millió évvel ezelőtt telepedett meg a szigeteken. Az ezt követő alkalmazkodó sugárzást a konkurencia hiánya és a sokféle üres fülke ösztönözte. Bár lehetséges, hogy egyetlen vemhes nőstény telepített be egy szigetet, valószínűbb, hogy egy fajból származó csoportról van szó.
A Hawaii-szigeteken más állatok és növények is vannak, amelyek hasonló, bár kevésbé látványos adaptációs sugárzásokon mentek keresztül.
Gyűrűs fajok
A biológiában a gyűrűs faj szomszédos populációk összefüggő sorozata, amelyek mindegyike képes kereszteződni a szomszédos populációkkal. A lánc két vége átfedésben van.
A sorozat két végpopulációja túlságosan távoli rokonságban áll ahhoz, hogy kereszteződhessenek. Az ilyen nem szaporodó, bár genetikailag kapcsolatban álló "végpopulációk" egyazon régióban együtt élhetnek, így zárva le egy "gyűrűt".
A gyűrűs fajok fontos bizonyítékai az evolúciónak: szemléltetik, hogy mi történik az idő múlásával, amikor a populációk genetikailag eltérnek egymástól. Richard Dawkins megfigyelte, hogy a gyűrűs fajok "csak a térbeli dimenzióban mutatnak meg nekünk valamit, aminek mindig az időbeli dimenzióban kell történnie".
Nehéz azonban egyszerű, egyértelmű példát találni.
Larus sirályok
A gyűrűs fajok klasszikus példája a Larus sirályok cirkumpoláris fajgyűrűje. E sirályok elterjedési területe egy gyűrűt alkot az Északi-sark körül, amelyet az egyes sirályok általában nem repülnek át.
Általánosságban elmondható, hogy a sirályok bizonyos mértékben kereszteződhetnek szomszédaikkal, kivéve a gyűrű két végén.
A kis fekete hátú sirályok és a hering sirályok eléggé különböznek egymástól ahhoz, hogy általában nem hibridizálódnak; így a sirályok csoportja kontinuumot alkot, kivéve ott, ahol a két vonal Európában találkozik.
Bár a valóság sokkal bonyolultabb, az általános elképzelést jól mutatja.
Ensatina szalamandrák
Az Ensatina szalamandra a kaliforniai Közép-völgy körüli hegyekben élő gyűrűs faj. A komplexum patkó alakot alkot a hegyek körül. Bár a patkó körüli 19 populáció mindegyike között előfordulhat kereszteződés, a patkó nyugati végén található Ensatina eschscholtzii alfaj nem kereszteződhet a keleti végén található Ensatina klauberi alfajjal. Ez a "fajképződési folyamat szinte minden szakaszának" (Dobzhansky) illusztrációja. Richard Highton azzal érvelt, hogy az Ensatina több faj esete, és nem egy faj folytonossága.
A zöldes pacsirta
A zöldike (Phylloscopus trochiloides) több alfajjal rendelkezik, amelyek közül a P. t . viridianus a legismertebb Európában. Ez egy gyűrűs faj, amelynek populációi a Tibeti-fennsíktól keletre és nyugatra térnek el egymástól, majd az északi oldalon találkoznak. Kapcsolatuk meglehetősen zavaros.




Szimpatrikus fajképződés
A szimpatrikus fajképződés azt jelenti, hogy egyetlen ősfajból két vagy több leszármazott faj jön létre, amelyek mind ugyanazt a földrajzi helyet foglalják el. Ezt ma már nagyon gyakorinak tartják.
A szimpatrikus fajmegújulás során a fajok ugyanazon a helyen élve térnek el egymástól. A szimpatrikus fajképződés gyakran említett példái azok a rovarok, amelyek ugyanazon a területen különböző gazdanövényektől válnak függővé.
A szimpatrikus fajképződés, mint a fajképződés mechanizmusának létezését hevesen vitatták. Az emberek azzal érveltek, hogy a szimpatrikus fajképződés bizonyítékai valójában a mikroföldrajzi fajképződés példái. Általánosságban elmondható, hogy ma már nem ez a preferált magyarázat. Inkább a közeli rokon fajok közötti hibridizáció eredménye, amelyet az ilyen keresztezések utódaira ható természetes szelekció követ. A szimpatrikus fajképződés egyik széles körben elfogadott példája a kelet-afrikai Nabugabo-tó cichlidái, amelyről úgy gondolják, hogy szexuális szelekciónak köszönhető.
Poliploidizáció révén történő fajmeghatározás
A poliploidia számos gyors fajkialakulási eseményt okozott, mivel például a tetraploid x diploid párosítások utódai gyakran triploid steril utódokat eredményeznek.
Nem minden poliploid azonban reproduktívan elszigetelt a szülői növényektől, és a génáramlás még mindig előfordulhat, például triploid hibrid x diploid párosításokon keresztül, amelyek tetraploidokat eredményeznek.
Úgy tűnik, hogy a létező növény- és állatfajok nagy része evolúciós története során poliploidizáción ment keresztül. A sikeres poliploid fajok szaporodása néha aszexuális, parthenogenezis útján történik. Ismeretlen okokból sok aszexuális szervezet poliploid.
Galagonya légy
Az evolúció működésének egyik példája a galagonyalégy, a Rhagoletis pomonella esete, amely a jelek szerint szimpatriás fajmegújuláson megy keresztül.
A galagonyalégy különböző populációi különböző gyümölcsökkel táplálkoznak. Egy különálló populáció a 19. században alakult ki Észak-Amerikában, nem sokkal azután, hogy az alma, amely nem őshonos faj, bekerült az országba. Ez az almával táplálkozó populáció általában csak almával táplálkozik, a galagonya történetileg kedvelt gyümölcsével nem. A jelenlegi galagonyatermő populáció általában nem táplálkozik almával.
Bizonyos bizonyítékok arra utalnak, hogy szimpatrikus fajképződés zajlik. A galagonyalégy a szezonban később érik, és tovább tart az ivarérés, mint az almalégyé; és kevés bizonyíték van a kereszteződésre (a kutatók 4-6%-os hibridizációs arányt dokumentáltak).
Az új galagonyalégy megjelenése a folyamatban lévő evolúció példája.
Hibridizáció
Ritkán új faj jön létre, amikor különböző fajok egyes tagjai párosodnak. Általában az ilyen hibrid párosítások termékei terméketlenek (nem termékenyek), vagy viszonylag terméketlenek, és így a természetes szelekció kiszorítja őket.
Egy új és sikeres hibrid fajra találtak példát. Az új faj az olasz és a spanyol veréb hibridje, és az alapvető tényeket a vérükben lévő DNS szekvenciaelemzésével ellenőrizték. A madár Olaszországban él, egy olyan régióban, ahol mindkét szülőfaj él. Nem szaporodik a spanyol verebekkel, annak ellenére, hogy egymás mellett él velük.
Mesterséges fajképződés
A háziasított állattartás révén új fajok jöttek létre, de a kezdeti időpontok és módszerek nem tisztázottak. A házi juhok például hibridizációval jöttek létre, és már nem hoznak létre életképes utódokat az Ovis orientalis fajjal, amelytől származnak.
A házi szarvasmarhák viszont a vadon élő ökör, gaur, jak stb. számos fajtájával azonos fajnak tekinthetők, mivel könnyen hozhatnak velük termékeny utódokat.
Laboratóriumi fajok
A legjobban dokumentált új fajok laboratóriumi létrehozása az 1980-as évek végén történt. William Rice és G. W. Salt gyümölcslegyeket, Drosophila melanogastereket tenyésztett ki, egy labirintus segítségével, amelyben három különböző élőhelyet - világos/sötét és nedves/száraz - választottak.
Minden egyes generációt a labirintusba helyeztek, és a nyolc kijárat közül kettőből két kijáraton kijövő legyek csoportjait elkülönítették, hogy a saját csoportjukban szaporodjanak egymással. Harmincöt generáció után a két csoport és utódaik szaporodási szempontból elszigetelődtek egymástól, mivel a legyek erősen preferálták az élőhelyeket: csak az általuk preferált területeken belül párosodtak, így nem párosodtak a más területeket preferáló legyekkel. Az ilyen kísérletek történetét Rice és Hostert (1993) írja le.
Diane Dodd azt is meg tudta mutatni, hogy a Drosophila pseudoobscura fajban a párzási preferenciákból hogyan alakulhat ki reproduktív izoláció mindössze nyolc generáció után, különböző tápláléktípusok, keményítő és maltóz felhasználásával.
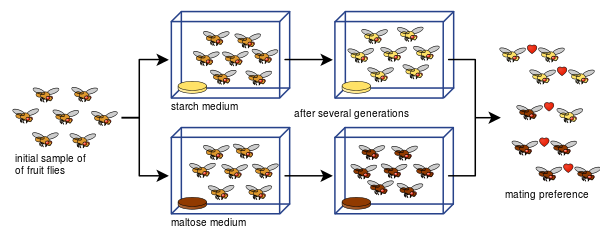
Dodd kísérletét sokan mások is könnyen megismételték, többek között más gyümölcslegyekkel és élelmiszerekkel.
Erősítés (Wallace-hatás)
A megerősítés az a folyamat, amelynek során a természetes szelekció növeli a reproduktív elszigeteltséget.
Előfordulhat, miután ugyanazon faj két populációja szétvált, majd ismét érintkezik egymással. Ha a szaporodási izolációjuk teljes volt, akkor már két különálló, összeférhetetlen fajjá fejlődtek.
Ha a szaporodási elszigeteltségük nem teljes, akkor a populációk közötti további párosodás hibrideket hoz létre, amelyek lehetnek termékenyek, de lehetnek nem termékenyek is. Ha a hibridek terméketlenek, vagy termékenyek, de kevésbé alkalmasak, mint az őseik, akkor nem lesz további szaporodási izoláció, és a fajképződés lényegében megtörtént (pl. mint a lovak és a szamarak esetében).
Ennek az az oka, hogy ha a hibrid utódok szülei mindegyike a saját, különböző környezetükhöz természetes módon kiválasztott tulajdonságokkal rendelkezik, a hibrid utódok mindkettőből hordozni fogják a tulajdonságokat, és nem illeszkednének egyik fülkébe sem olyan jól, mint bármelyik szülő. A hibridek alacsony fitneszértéke miatt a szelekció az asszortatív párosodásnak kedvezne, ami csökkentené a hibridizáció mértékét.
Ezt néha Wallace-hatásnak nevezik Alfred Russel Wallace evolúcióbiológus után, aki a 19. század végén felvetette, hogy ez fontos tényező lehet a fajképződésben.
Ha a hibrid utódok alkalmasabbak, mint az őseik, akkor a populációk visszaolvadnak ugyanabba a fajba azon a területen, amellyel érintkeznek.
Megerősítésre akkor van szükség az elkülönítéshez, ha egy faj két formája között "hibrid zóna" van. A hibrid zónák olyan területek, ahol az egymástól eltérő populációk találkoznak és kereszteződnek. A hibrid utódok nagyon gyakoriak ezekben a régiókban, amelyeket általában az egymástól eltérő fajok másodlagos érintkezése hoz létre.
Megerősítés nélkül a két populáció vagy faj kontrollálhatatlanul keveredne egymással. A megerősítést mesterséges szelekciós kísérletekben lehet előidézni a fent leírtak szerint.
Kérdések és válaszok
K: Mi az a fajmeghatározás?
V: A fajképződés a fajok kialakulásának folyamata. Ez az evolúciós biológia egyik legfontosabb része.
K: Mit gondolt Darwin a fajok kialakulásáról?
V: Darwin úgy gondolta, hogy a legtöbb faj közvetlenül a már létező fajokból származik, amit ő anagenezisnek vagy "phyletikus evolúciónak" nevezett.
K: Hogyan tekintettek általában a fajképződésre a 20. században?
V: A 20. században a tudósok úgy gondolták, hogy a legtöbb faj korábbi fajok szétválásával jött létre, amit kladogenezisnek neveztek. Azt is hitték, hogy ezt a szétválást olyan elkülönítő mechanizmusok okozták vagy segítették elő, mint például a fizikai elkülönülés.
K: Hogyan változtatták meg a legújabb kutatások a fajképződéssel kapcsolatos ismereteinket?
V: A legújabb kutatások kimutatták, hogy a rokon fajok között hibridizáció történhet, és a gének átadhatók ezeken a kereszteződéseken keresztül. Ez azt jelenti, hogy a reproduktív izoláció nem mindig szükséges egy faj meghatározásához, és az allopatria (fizikai elkülönülés) nem feltétlenül szükséges a fajképződéshez.
K: Mi köze van a fizikai elkülönülésnek az új fajok kialakulásához?
V: Az egykor egymás mellett létező fajok közötti fizikai elkülönülést tartják az új fajok kialakulásának egyik fő tényezőjének, amit számos, a tudományos szakirodalomban tárgyalt példa szemléltet.
Kapcsolódó cikkek
Szerző
AlegsaOnline.com Fajképződés (speciáció): meghatározás, mechanizmusok és példák Leandro Alegsa
URL: https://hu.alegsaonline.com/art/92570
Források
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833